ФОТОСИНТЕЗ
Расстановка ударений: ФОТОСИ`НТЕЗ
ФОТОСИНТЕЗ (от греч. phos, род. падеж photos — свет и synthesis — соединение), превращение зелёными растениями и фотосинтезирующими микроорганизмами лучистой энергии Солнца в энергию химич. связей органич. в-в. Единств. биол. процесс, к-рый идёт с увеличением свободной энергии и прямо или косвенно обеспечивает доступной химич. энергией все организмы (кроме хемосинтезирующих). Ежегодно в результате Ф. на Земле образуется ок. 150 млрд. т органич. в-ва. При этом усваивается ок. 300 млрд. т СO2 и выделяется ок. 200 млрд. т свободного O2.
Ф. высших растений осуществляется в хлоропластах, содержащих фотосинтетич. пигменты (хлорофиллы, каротиноиды). В основе этого сложного многоступенчатого процесса, протекающего как на свету (световая стадия Ф.), так и в темноте (темновая стадия Ф.), лежат окислительно-восстановит. реакции, в к-рых донором водорода и источником выделяемого кислорода служит H2О, а источником углерода — СO2. Лист поглощает ок. 90% падающей на него радиации в области спектра 400—700 нм (т. н. фотосинтетически активная радиация, или ФАР). Световая стадия Ф. начинается с улавливания хлоропластами ФАР. Свет возбуждает молекулы хлорофиллов. Возникает поток электронов, происходит фотодиссоциация воды и восстановление никотинамидаденинди-нуклеотидфосфата (НАДФ+) водородом воды до НАДФ х Н2. Свободный кислород, освобождаемый из воды, выделяется из листа. Сопряжённо с этим идёт фотофосфорилирование: образование богатого энергией соединения — аденозинтрифосфата (АТФ) из аденозиндифосфата и минер. фосфата за счёт энергии квантов света. АТФ и НАДФ х Н2 — осн. продукты, в к-рых запасается энергия света в ходе окислительно-восстановит. реакций Ф.; они используются затем в темновых процессах восстановления СO2, поступающего в межклетники листа, клетки и хлоропласты за счёт диффузии (диффузионная стадия Ф.). Во время темновой стадии Ф. происходит ферментативное присоединение поглощённой СO2 к органич. субстрату — рибулозодифосфату (РДФ). Образованное в-во включается в цикл последовательных ферментативных реакций (цикл Калвина), в результате к-рых при помощи НАДФ х Н2 и АТФ образуется молекула углевода (глюкозы). Различают растения, у к-рых присоединение СO2 к РДФ происходит непосредственно (С3-растения) или после предварит. связывания и образования органич. к-т на свету (С4-растения) или в темноте (толстянковые). Эффективность использования света на Ф. (кпд) мала. Большая часть поглощённой листом энергии теряется на тепловое излучение. В энергию химич. связей включается в ср. 1—2% поглощённой ФАР.
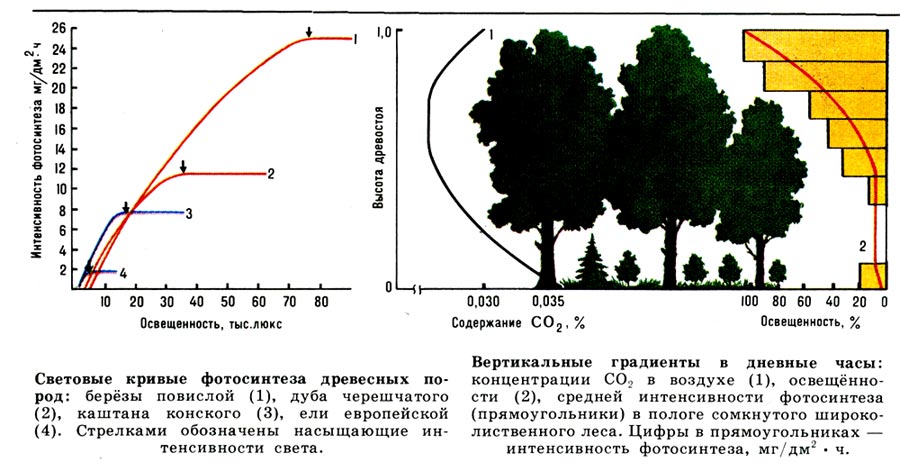
Все реакции Ф. находятся под влиянием внутр. и внеш. факторов (интенсивности света, содержания СO2 в воздухе, темп-ры воздуха, условий питания, оводнённости растений и т. д.), к-рые влияют на продуктивность Ф. Измерение Ф. чаще всего производится путём определения кол-ва поглощённой листом СO2 или выделенного O2. Одновременно с Ф. в листьях идёт процесс дыхания, при к-ром СO2 выделяется, а O2 поглощается. Поэтому различают наблюдаемый Ф. (нетто-Ф.), или газообмен, т. е. кол-во газа, выделившегося или поглощённого в результате двух противоположно направленных процессов, и истинный Ф. (брутто-, или грос-Ф., при к-ром дыхание исключено). Осн. показатель Ф.— его интенсивность, т. е. кол-во газа, поглощённого или выделенного единицей массы или поверхности листа в единицу времени. Интенсивность Ф. зависит от вида растений, состояния листьев, внеш. условий (свет, СO2). Ф. лесных древесных растений в 5—8 раз ниже, чем Ф. травянистых растений открытых местообитаний. Среднедневная интенсивность Ф. взрослых листьев листопадных деревьев равна 10—20 мг СO2/дм2- ч, вечнозелёных хвойных — 4—15 мг СO2/дм2- ч. Поглощение СO2 листьями начинается по достижении ими 1/4— 1/3 окончат. размера. В начале роста, когда Ф. очень слабый, а дыхание усилено, листья только выделяют СO2. С ростом листа интенсивность Ф. увеличивается и становится максимальной у листьев, достигших 3/4 конечного размера, затем несколько снижается и резко уменьшается в период пожелтения листьев. С увеличением освещённости интенсивность Ф. возрастает, пока не достигнет насыщения (уровня плато, или Фмакс). Насыщающая интенсивность света и уровень плато Ф. у разных видов растений различны и определяются степенью их светолюбия, световыми условиями выращивания и др. факторами. Светолюбивые виды растений (берёза, лиственница, сосна) наиболее эффективно используют для Ф. свет высокой интенсивности, а теневыносливые (ель, пихта, бук)— низкой. Теневые листья содержат больше хлорофилла, чем световые. Степень изменения Ф. у одного и того же вида растений под влиянием световых условий выращивания определяется лабильностью фотосинтетич. аппарата (анатомич. структурой, содержанием хлорофилла и пр.), что обусловливает возможность адаптации фотосинтетич. аппарата к условиям освещения. Фмакс у световых и теневых листьев клёна остролистного различается в 9 раз, у берёзы повислой — в 4 раза, у осины — в 1,5 раза.
При увеличении содержания СО2 в воздухе Ф. увеличивается. Углекислотное насыщение Ф. достигается при содержании СO2 приблизительно в 3 раза выше нормального её содержания в атмосфере (0,03%). Миним. концентрация СO2, при к-рой начинается её поглощение листьями, 0,003—0,004%. Действие темп-ры на Ф. связано с её влиянием на темновые реакции и вторичные процессы. Поглощение СO2 с повышением темп-ры ускоряется, пока не будет достигнут нек-рый оптим. уровень, к-рый сохраняется в довольно широком диапазоне темп-р (15—30°). Он зависит от вида растений, сезона, климатич. зон. Минимальная, прекращающая Ф. темп-pa для хвойных —6°, для листв. пород —1°, максимальная — соответственно 30—35° и 40—45°.
Продолжительность Ф. листопадных пород в умеренном поясе ок. 130 дней, вечнозелёных хвойных — на 30—40 дней больше. Ф. хвои прошлых лет ниже, чем Ф. закончившей рост молодой хвои. Дневная динамика Ф. под влиянием внеш. факторов может иметь форму одно-, двух- или многовершинной кривой. В зоне умеренного климата продолжительность Ф. световых листьев (верх. полог, опушка) в летние месяцы равна 15 ч, теневых листьев (ниж. полог) — 8—10 ч. Градиент изменений факторов внеш. среды под пологом сомкнутого древостоя обусловливает закономерные различия ср. интенсивности Ф. по высоте полога. Так, закономерное уменьшение освещённости и содержания СO2 по мере углубления внутрь полога фитоценоза приводит к снижению интенсивности Ф. В самом ниж. ярусе лесного фитоценоза интенсивность Ф. вновь слегка увеличивается за счёт повышенного содержания СO2 в приземном слое воздуха.
Данные о ср. интенсивности Ф., времени работы и массе листьев на 1 га древостоя (см. Индекс листовой поверхности) используют для расчёта газообмена леса с окружающей атмосферой и для оценки первичной биологической продуктивности. В высокопроизводительных сомкнутых древостоях умеренного климата лиственные или хвойные породы способны за вегетац. сезон поглотить 20—25 т/га СO2 и выделить 15—18 т/га O2. При этом создаётся 14—18 т/га органич. в-ва. В дождевых тропич. лесах поглощение СO2 за год достигает ок. 150 т/га, а выделение O2 — ок. 110 т/га. Первичная биол. продукция — ок. 40 т/га. Неблагоприятные почвенные условия (недостаток питат. в-в, переувлажнение или недостаток влаги, наличие токсич. в-в, переуплотнение почвы) ведут к снижению как интенсивности Ф., так и продуктивности леса. Повышение продуктивности при этом возможно путём мелиорации земель. В перегущенных насаждениях, где интенсивность Ф. снижена из-за недостатка света, повышение Ф. достигается за счёт своевременных рубок ухода. Биол. продуктивность леса при этом не повышается, но увеличивается выход хозяйственно ценной древесины.
(Цельникер Ю. Л., Физиологические основы теневыносливости древесных растений, М., 1978; Белл Л. Н., Энергетика фотосинтезирующей растительной клетки, М., 1980; Лир X., Польстер Г., Фидлер Г. И., Физиология древесных растений, пер. с нем., М., 1974; Лархер В., Экология растений, пер. с нем., М., 1978; Xолл Д., Рао К. В., Фотосинтез, пер. с англ., М., 1983.)
Источники:
- Лесная энциклопедия: В 2-х т., т.2/Гл.ред. Воробьев Г.И.; Ред.кол.: Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. энциклопедия, 1986.-631 с., ил.
|
ПОИСК:
|
При копировании материалов проекта обязательно ставить активную ссылку на страницу источник:
http://dendrology.ru/ 'Книги о лесе и лесоводстве'